Gametogeneza

Spis treści
Jak powstają komórki jajowe i plemniki ?
Nasz materiał genetyczny, czyli DNA (kwas deoksyrybonukleinowy) znajduje się w każdej komórce w jądrze komórkowym. Dzięki temu komórka posiada wszystkie niezbędne dla niej informacje związane z jej wzrostem, funkcjonowaniem i reakcją na bodźce, czynniki środowiskowe. Taki zestaw komórkowego DNA nazywamy genomem. DNA nie występuje w komórce samodzielnie, lecz jest połączone ze specjalnymi białkami, tzw. histonami, które nadają mu odpowiednią strukturę. W dużym uproszczeniu możemy powiedzieć, że nici DNA są nawinięte na histony jak na szpulkę tworząc tzw. nukleosomy (inaczej nici nukleosomowe, włókna nukleosomowe). W jądrze komórkowym oprócz DNA i histonów znajdują się jeszcze białka niehistonowe oraz niewielkie ilości RNA (kwasu rybonukleinowego) – wszystkie one razem tworzą chromatynę. Przez większą część życia komórki chromatyna znajduje się w tzw. postaci zdekondensowanej, czyli występuje w postaci długich, cienkich nici z zawijasami – nukleosomów. Dzięki temu wszystkie informacje zapisane w genach są „łatwo” dostępne dla białek, które je odczytują. Sytuacją, w której chromatyna ulega kondensacji jest podział komórki. Wtedy to możemy zauważyć, że chromatyna nie jest jedną długą nicią, lecz jest ona podzielona na odcinki. Każdy taki odcinek chromatyny dopiero po kondensacji, czyli takim zagęszczeniu, „ścieśnieniu się” przyjmuje charakterystyczny kształt litery X, którą mają chromosomy.
Reasumując – chromosomy w swojej skondensowanej formie ściśle upakowanej chromatyny takiej, jaką znamy ze zdjęć mikroskopowych i rysunków są widoczne w komórce tylko w czasie kiedy komórka się dzieli. W pozostałym okresie cyklu komórkowego chromosomy o kształcie litery X nie są dostrzegalne, gdyż chromatyna jest wtedy zdekondensowana, możemy potocznie powiedzieć, że nić nukleosomowa jest wtedy rozluźniona, rozpleciona.
Każdy gatunek ma swoją własną, charakterystyczną liczbę chromosomów. Na przykład koty mają 38 chromosomów (19 par), psy 78 chromosomów (39 par), owce mają 54 chromosomy (27 par), a ludzie 46 chromosomów (23 pary). Jak wiele gatunków, ludzie są diploidalni, co oznacza, że ich chromosomy połączone są w pary, czyli 46 ludzkich chromosomów tworzy 23 pary. Komórki diploidalne oznaczamy na schematach jako 2n.
W tym 22 pary to tzw. chromosomy autosomalne, gdzie każdy chromosom pary jest homologiem drugiego chromosomu z pary. Chromosomy w parze nazywane są chromosomami homologicznymi gdyż są do siebie bardzo podobne. Mają ten sam kształt i rozmiar oraz znajdują się na nich informacje genetyczne dotyczące tej samej cechy, funkcji, białka itp. Innymi słowy mają te same geny, których położenie na każdym z chromosomów homologicznych jest też takie samo. Ale co ważne – ten sam gen nie oznacza, że zapisana informacja na obu chromosomach z pary jest taka sama. Pamiętajmy, że jeden chromosom z pary to ten odziedziczony po matce, a drugi po ojcu. A więc możemy mieć różne warianty tego samego genu. Przykładem może być grupa krwi – na jednym chromosomie homologicznym możemy mieć zapisaną grupę krwi A, a na drugim chromosomie z tej pary grupę krwi B (mamy wtedy grupę krwi AB). Oba chromosomy zawierają więc informację genetyczną dotyczącą tej samej cechy – grupy krwi, ale geny nie są jednakowe, tylko występują w 2 różnych wariantach – A i B. Może oczywiście być też tak, że mamy ten sam wariant genu na obu chromosomach, np. grupę krwi B.
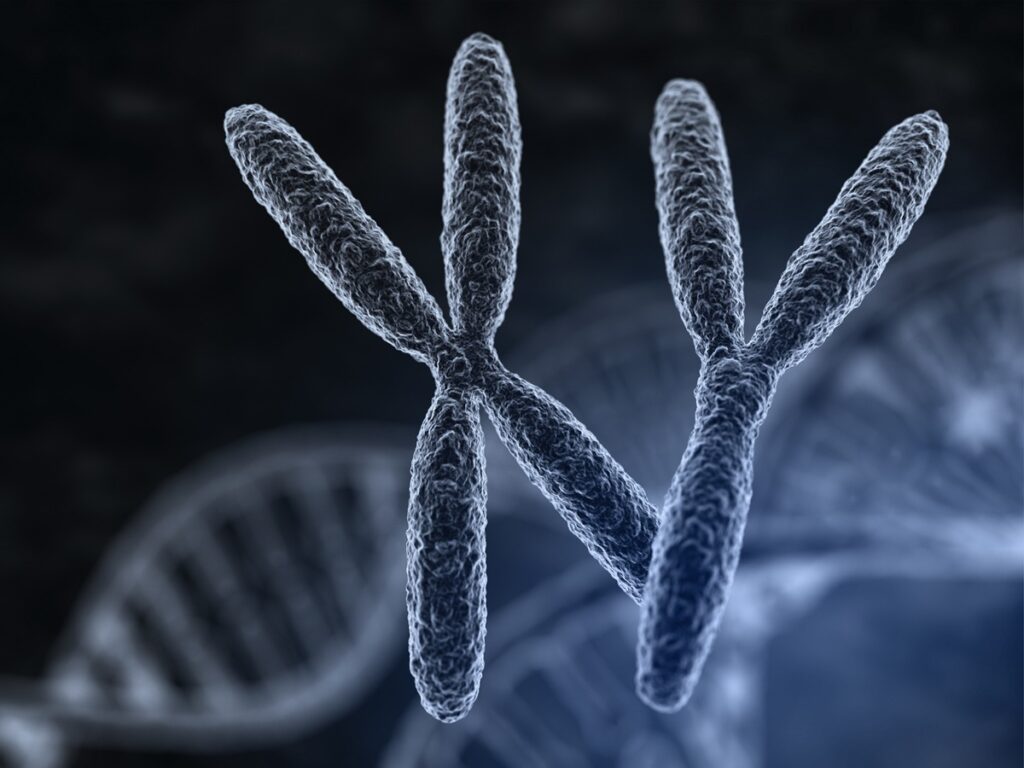
Natomiast 23. para chromosomów stanowi wyjątek – te chromosomy nie są prawdziwymi homologami. Jest to para chromosomów płciowych (inaczej allosomalnych), w przypadku kobiety mamy 2 chromosomy X (XX), a w przypadku mężczyzny 1 chromosom X i 1 chromosom Y (XY). W tych chromosomach tylko niewielka część to regiony podobne, na których znajdują się te same geny (są to geny związane z rozmnażaniem i powstawaniem, dzieleniem się komórek płciowych). Poza tymi genami pozostałe obszary informacji genetycznej na chromosomach X i Y są różne, kodują one tzw. cechy sprzężone z płcią (np. hemofilia, daltonizm).
Takie zestawy 23 par chromosomów znajdują się w każdej komórce ludzkiego organizmu. Wyjątek stanowią gamety płciowe (komórki jajowe i plemniki), które są haploidalne (oznaczamy je na schematach jako n) i mają po 23 chromosomy (po jednym chromosomie z każdej pary). Dzięki temu po zapłodnieniu (połączeniu się komórki jajowej z plemnikiem) powstaje zygota, która jest już komórką diploidalną i z niej po kolejnych podziałach komórkowych powstanie nowy organizm. Zygota posiada połączony z obu komórek płciowych materiał genetyczny (połowa od matki, połowa od ojca).
W jaki sposób dzielą się komórki?
Aby nasz organizm mógł rosnąć, rozwijać się i regenerować jego komórki muszą ulegać podziałom, czyli muszą powstawać nowe komórki.
Komórka, aby mogła się podzielić musi przejść przez kolejne etapy:
- musi urosnąć
- musi powielić (zduplikować, podwoić) swój materiał genetyczny, aby połowę przekazać kolejnej komórce
- musi fizycznie podzielić się na 2 komórki potomne
Wszystkie te procesy to seria dobrze zorganizowanych etapów zachodzących po sobie, czyli tzw. cykl komórkowy. Co więcej, wszystkie procesy w cyklu komórkowym nieustannie się powtarzają, zawsze w tej samej, określonej kolejności, czyli powstałe po podziale komórki potomne rosną, powielają swój materiał genetyczny i znów się dzielą. W komórkach eukariotycznych (czyli posiadających jądro komórkowe), cykl komórkowy składa się z dwóch faz głównych: interfazy i mitozy.
- podczas interfazy komórka przygotowuje się do podziału – rośnie i kopiuje swój materiał genetyczny (DNA)
- podczas mitozy następuje podział komórki (jej cytoplazmy, organelli komórkowych i DNA na dwa zestawy tworząc dwie nowe komórki)
Interfaza, czyli przygotowanie do podziału odbywa się w 3 etapach:
- faza G1 – następuje wzrost masy i objętości komórki, intensywna synteza białek strukturalnych i regulatorowych
- faza S – komórka produkuje w swoim jądrze pełną kopię DNA (podwaja materiał genetyczny), czyli następuje tzw. replikacja genomu oraz zachodzi synteza histonów
- faza G2 – komórka dalej rośnie, produkuje białka wrzeciona podziałowego, białka niehistonowe (niezbędne w kondensacji chromatyny), białka i lipidy niezbędne do odtworzenia błony komórkowej oraz następuje podział organelli samoreplikujących się (mitochondria)
Gdy komórka jest gotowa do podziału kończy się faza G2 i rozpoczyna się mitoza, która składa się z 2 procesów:
- kariokinezy – podział jądra komórkowego (materiału genetycznego)
- cytokinezy – podział cytoplazmy
W wyniku tych procesów z każdej komórki powstają dwie komórki potomne identyczne genetycznie.
Jak dokładnie przebiega mitoza?
Profaza – jest to pierwsza faza podziału mitotycznego komórki. Pamiętamy, że komórka przygotowała się już do podziału – ma podwójną ilość DNA (oznaczamy to jako 4c DNA). Chromatyna zaczyna się kondensować i tworzy pałeczkowate struktury w kształcie litery X zwane chromosomami. Chromosomy składają się z dwóch jednakowych części, czyli chromatyd siostrzanych, które połączone są ze sobą centromerem. Możemy je zaobserwować tylko w trakcie podziału komórki, kiedy następuje kondensacja podwojonej ilości DNA. W tej fazie rozpoczyna się również tworzenie wrzeciona kariokinetycznego (podziałowego), czyli takiej specjalnej struktury zbudowanej z mikrotubul, która ułatwi prawidłowy podział materiału genetycznego. Na każdej chromatydzie siostrzanej powstają kinetochory, czyli miejsca łączenia się mikrotubul wrzeciona podziałowego z chromatydą.

Metafaza – mikrotubule wrzeciona podziałowego łączą się z kinetochorami, „spinając” chromosomy z biegunami komórki i chromosomy ustawiają się w centralnej części komórki w płaszczyźnie równikowej tworząc tzw. płytkę metafazową.

Anafaza – na chromatydy siostrzane zaczynają oddziaływać siły mające na celu oderwanie ich od siebie, centromery chromosomów ulegają podziałowi. Dzięki temu chromatydy siostrzane (dwie identyczne połowy zduplikowanego chromosomu) odsuwając się od siebie, stają się dwoma oddzielnymi chromosomami potomnymi. Skracają się mikrotubule wrzeciona, dzięki czemu chromosomy potomne przemieszczają się do przeciwległych biegunów komórki. Moment, gdy centromery chromosomów są od siebie maksymalnie oddalone uważany jest za koniec anafazy. Pod koniec tej fazy stopniowo zanika też wrzeciono podziałowe. Pod koniec anafazy rozpoczyna się podział cytoplazmy i znajdujących się w niej organelli na 2 równe części. Podział ten zachodzi przy udziale wrzeciona cytokinetycznego.
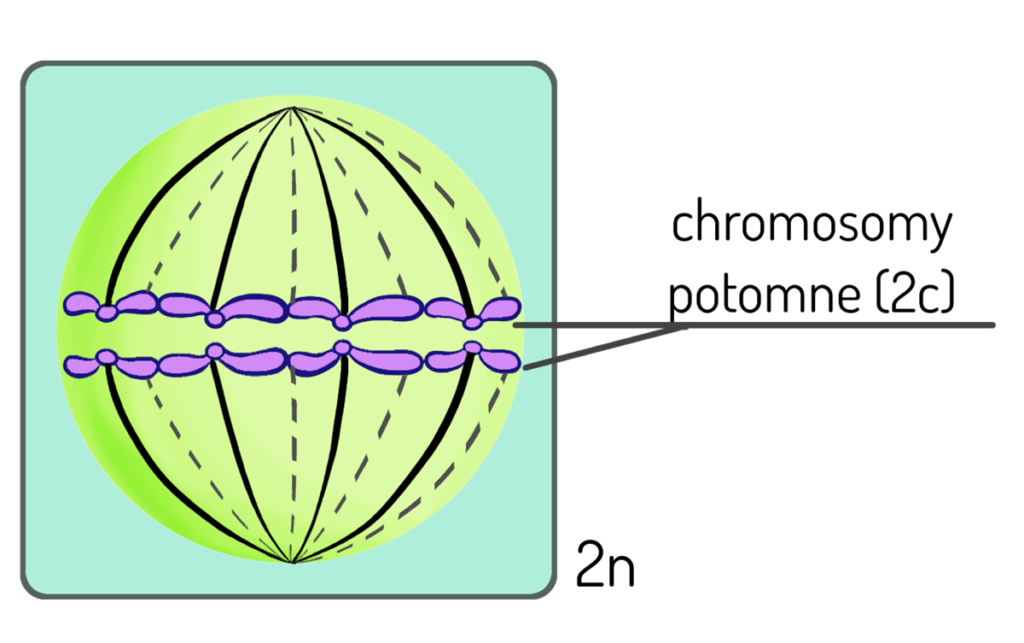
Telofaza – chromosomy potomne osiągnęły maksymalny stopień oddalenia i znajdują się na dwóch przeciwległych biegunach komórki. Odtwarzają się błona jądrowa i jąderko, które zaniknęły pod koniec profazy. Chromosomy ulegają dekondensacji i znowu tworzą siateczkę chromatynową. W komórce powstają 2 jądra potomne o takiej samej liczbie chromosomów i takiej samej informacji genetycznej, co ulegająca podziałowi komórka macierzysta. Dochodzi do ostatecznego podziału cytoplazmy i odtwarza się błona komórkowa, która dzieli komórkę macierzystą na 2 diploidalne komórki potomne (2 x 2n).

Komórki organizmu wielokomórkowego jakim jest człowiek znajdują się w różnych fazach cyklu komórkowego. W zależności od rodzaju komórek, czas trwania cyklu komórkowego różni się. Jedne komórki dzielą się szybciej, inne wolniej. Co więcej, jedne komórki potomne ulegają dalszym podziałom, a inne po podziale przechodzą w tzw. fazę spoczynku (fazę G0). Na przykład komórki szpiku kostnego lub komórki nabłonkowe jelita są stale w cyklu komórkowym i ulegają ciągłym podziałom. Z kolei całkowicie zróżnicowane neurony, kardiomiocyty, hepatocyty znajdują się w fazie G0 i nie przygotowują się do podziału, tylko wykonują swoje zadania (np. przewodzą sygnały, kurczą się, magazynują węglowodany itp.).
Jak powstają gamety płciowe?
Wiemy już w jaki sposób powstają, dzielą się praktycznie wszystkie komórki naszego organizmu. Wyjątek stanowi powstawanie komórek rozrodczych (gamet płciowych), czyli plemników i komórek jajowych.
Wszystkie komórki naszego ciała za wyjątkiem komórek płciowych to tzw. komórki somatyczne. Wiemy już, że komórki te są diploidalne (2n) i mają zestaw 23 par chromosomów (razem 46 chromosomów), natomiast komórki rozrodcze są haploidalne (n) i posiadają 23 chromosomy, czyli połowę tego co ma komórka macierzysta i każda komórka somatyczna. Jak więc dochodzi do ich powstania? Proces ten jest bardziej skomplikowany niż mitoza, gdyż przeprowadza nas od komórki diploidalnej (tej z dwoma zestawami chromosomów), do komórek haploidalnych (tych z pojedynczym zestawem chromosomów) i nazywany jest mejozą. W mejozie pomimo, że poszczególne etapy i sposób organizacji podziału materiału genetycznego jest podobny jak w procesie mitozy, to jednak zadanie jest trudniejsze, gdyż w trakcie podziału muszą rozdzielić się nie tylko chromatydy siostrzane, ale również chromosomy homologiczne (czyli chromosomy jednej pary, które otrzymaliśmy jeden od matki, a drugi od ojca). Dlatego też mejoza jest dwustopniowym procesem podziału. Pary chromosomów homologicznych rozdzielają się podczas pierwszej części podziału komórki, nazywanej mejozą I. Chromatydy siostrzane oddzielają się podczas drugiej części podziału, nazywanej mejozą II. W każdym z tych 2 etapów mejozy komórki przechodzą przez cztery fazy: profazę, metafazę, anafazę i telofazę (takie jak w mitozie).
Zanim rozpocznie się mejoza, komórka na początku musi przejść przez interfazę. Dokładnie tak jak w mitozie, komórka najpierw rośnie w fazie G1, duplikuje swój materiał genetyczny (wszystkie swoje chromosomy) podczas fazy S, a następnie przygotowuje się do podziału podczas fazy G2.
Mejoza I – rozdział chromosomów homologicznych
Profaza I – tak jak w mitozie następuje kondensacja chromatyny, ale oprócz tego chromosomy homologiczne układają się w pary. Każdy chromosom dokładnie układa się wraz ze swoim homologicznym partnerem i oba dopasowują się odpowiadającymi między sobą pozycjami na chromosomach na całej ich długości. Wtedy zachodzi bardzo charakterystyczne dla mejozy zjawisko, tzw. crossing over. Polega ono na tym, że pomiędzy chromosomami homologicznymi dochodzi do wymiany odcinków chromatyd. Innymi słowy następuje wymiana materiału genetycznego pomiędzy chromosomami tej samej pary. Dzięki temu materiał genetyczny od matki i od ojca zostaje „wymieszany”. Zapewnia tą większą różnorodność w obrębie naszego gatunku. Crossing over jest jednym z elementów rekombinacji genetycznej, czyli właśnie procesów wymiany materiału genetycznego, w wyniku których powstają nowe genotypy. Rekombinacja genetyczna stanowi główne źródło zmienności dziedzicznej przyrodzie, jest też podstawą dla procesów ewolucyjnych. Warto pamiętać, że rekombinacja genetyczna, a w tym crossing over nie prowadzą do powstawania nowych genów, lecz przyczyniają się do tworzenia różnych kombinacji już istniejących wariantów genów. Oprócz crossing over, rekombinacja genetyczna jest wynikiem losowego łączenia się gamet płciowych w trakcie zapłodnienia oraz losowego rozdziału chromosomów homologicznych w trakcie mejozy I. Na poniższym rysunku schematycznie przedstawiony jest proces crossing over (w rzeczywistości chromosomy homologiczne są położone jeden na drugim i w niektórych miejscach ich chromatydy są połączone – miejsca te nazywamy chiazmami, na schemacie pokazaliśmy je obok siebie, aby lepiej zobrazować crossing over).

Metafaza I – po crossing over, wrzeciono kariokinetyczne wyłapuje pary chromosomów homologicznych (nazywamy je biwalentami) i ustawia je w płaszczyźnie równikowej w centrum komórki. Czyli w przypadku człowieka – komórka ma 46 chromosomów, a z nich powstają 23 biwalenty. Biwalenty ułożone w linii tworzą płytkę metafazową. I to jest kolejna różnica pomiędzy mejozą a mitozą – w metafazie I, to pary chromosomów homologicznych, a nie indywidualne chromosomy, układają się do rozdziału w linię w płytce metafazowej.
Anafaza I – każdy chromosom z pary chromosomów homologicznych przemieszcza się przy udziale wrzeciona kariokinetycznego do przeciwległego bieguna komórki. Ale w tym czasie chromatydy siostrzane każdego chromosomu pozostają przyłączone jedna do drugiej i nie rozdzielają się.
Telofaza I – każdy chromosom z pary znajduje się po przeciwległej stronie komórki i tworzą się 2 jądra potomne (każde ma po 23 chromosomy z dwoma chromatydami siostrzanymi). W pierwszym podziale mejotycznym może, ale nie musi zachodzić proces cytokinezy. Zazwyczaj niemal bezpośrednio po pierwszym podziale mejotycznym zachodzi drugi podział mejotyczny (mejoza II).

Mejoza II – rozdział chromatyd siostrzanych
Komórki, które powstały po pierwszym podziale mejotycznym to już komórki haploidalne – mają tylko jeden chromosom z każdej homologicznej pary, ale ich chromosomy nadal składają się z dwóch chromatyd siostrzanych. Czyli mamy pojedyncze chromosomy, ale z podwójną ilością DNA (podwojenie materiału genetycznego miało miejsce w interfazie przed mejozą I). Mejozy II nie poprzedza już interfaza, a więc komórki przechodzą od mejozy I do mejozy II już bez kopiowania swojego DNA. Dzięki temu podczas mejozy II chromatydy siostrzane rozdzielają się i powstają komórki haploidalne z niezduplikowanymi chromosomami, czyli nasze gamety płciowe. Dzięki temu po połączeniu się podczas zapłodnienia żeńskiej i męskiej gamety płciowej w zygotę powstaje jedna komórka diploidalna, czyli taka, która ma 23 pary chromosomów homologicznych. Mejoza II jest już procesem krótszym i prostszym niż mejoza I, w jej przebiegu zachowane są takie same 4 etapy jak w procesie mitozy (profaza, metafaza, anafaza i telofaza).
Ostatecznie w wyniku obu podziałów mejotycznych z macierzystej komórki płciowej powstają 4 komórki haploidalne.

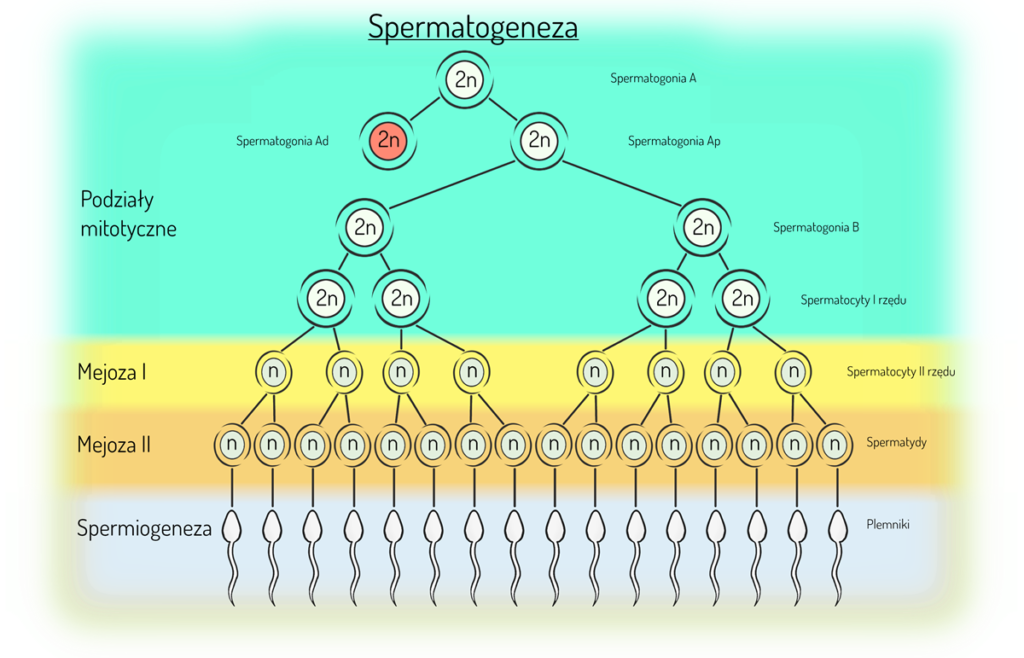
Proces powstawania plemników to spermatogeneza, a komórek jajowych to oogeneza.
Spermatogeneza
Spermatogeneza zachodzi w gonadach męskich czyli jądrach.
Rozpoczyna się w okresie pokwitania i ma 3 główne fazy:
- Spermatogoniogeneza – diploidalne komórki macierzyste (spermatogonia typu A) dzielą się mitotycznie, dzięki czemu powstają kolejne komórki stanowiące taką bazę, źródło komórek, z których w przyszłości wykształcą się plemniki. Następnie losy spermatogonii typu A mogą przebiegać dwutorowo. Jedna część (tzw. spermatogonia Ad, czyli ciemne od ang. dark) będzie stanowiła rezerwuar dla nowych generacji spermatocytów typu A, a druga część (tzw. spermatogonia Ap, czyli jasne od ang. pale) również dzieląc się mitotycznie przekształci się w spermatogonia typu B. Ze spermatogoniów typu B po kolejnych podziałach mitotycznych utworzą się spermatocyty I rzędu (dalej są to komórki diploidalne).
- Spermatocytogeneza – spermatocyty I rzędu przechodzą już podziały mejotyczne. W wyniku pierwszego podziału mejotycznego powstaną spermatocyty II rzędu (komórki haploidalne, ale każdy chromosom ma 2 chromatydy siostrzane, czyli podwojoną ilość DNA). Na drodze drugiego podziału mejotycznego spermatocyty II rzędu podzielą się na spermatydy, które są komórkami haploidalnymi z pojedynczą ilością DNA. Z każdego spermatocytu I rzędu powstaną 4 spermatydy, a więc finalnie 4 plemniki.
- Spermiogeneza – spermatydy przeobrażają się w plemniki.
Cykl spermatogenezy trwa u człowieka ok. 74 dni, w tym czas wzrostu, dojrzewania i podziałów poszczególnych komórek wynosi średnio:
- spermatogonia Ap – 18 dni
- spermatogonia B – 9 dni
- spermatocyty I rzędu – 23 dni
- spermatocyty II rzędu – 1 dzień
- spermatydy – 23 dni
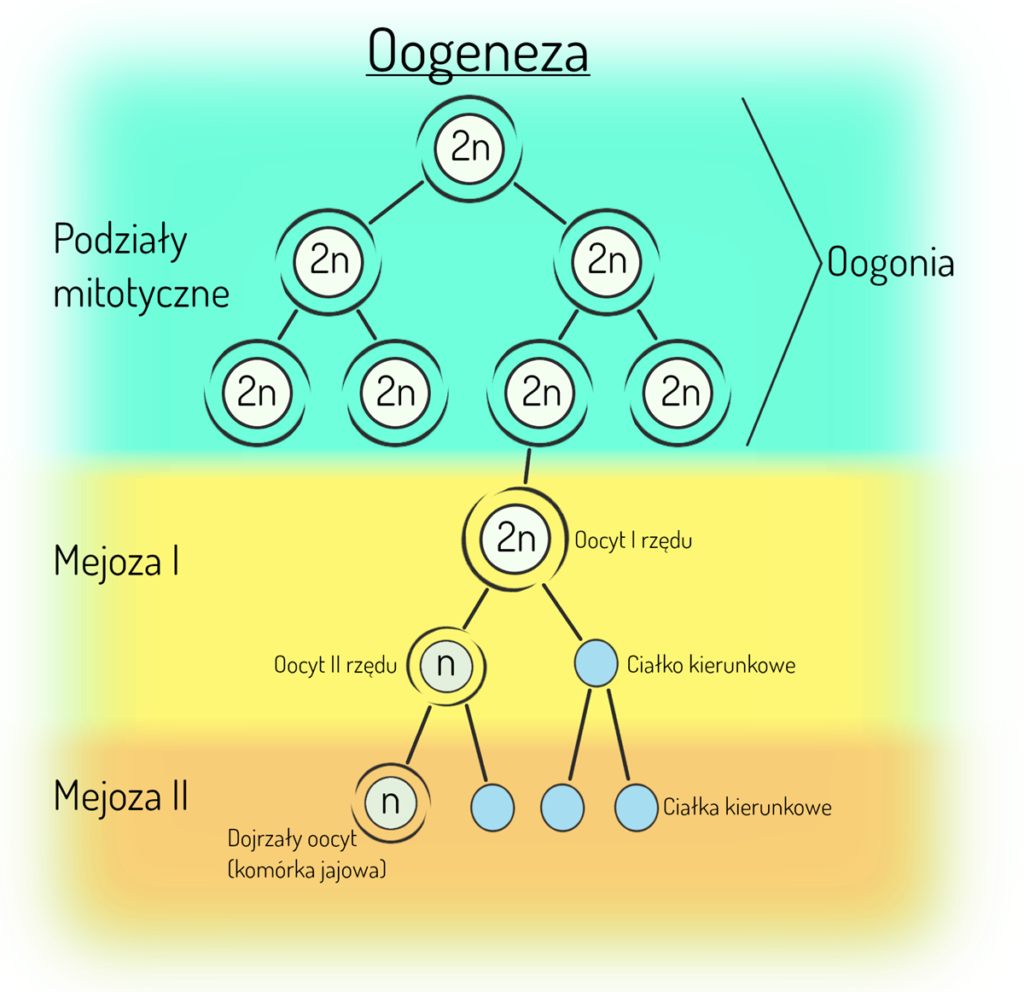
Oogeneza
Oogeneza, czyli proces powstawania i dojrzewania gamet żeńskich – komórek jajowych zachodzi w gonadach żeńskich, czyli w jajnikach. Oogeneza różni się od spermatogenezy m.in. rozłożeniem w czasie poszczególnych etapów podziałów i dojrzewania komórek rozrodczych – znaczna część oogenezy zachodzi już w trakcie życia płodowego. Żeńskie pierwotne komórki płciowe, tzw. gonocyty pojawiają się w zarodku ludzkim już w ścianie pęcherzyka żółtkowego. Ponieważ mają zdolność ruchu, stąd przemieszczają się do zawiązków gonad. Wnikając do nich są sygnałem do różnicowania się zawiązków w jajniki. Od tego momentu gonocyty nazywane są oogoniami (lub oocytami).
- W okresie pomiędzy 10. a 32. tygodniem ciąży oogonia namnażają się przez podział mitotyczny (są one komórkami diploidalnymi).
- Badania wykazują, że niewielka część oogonii już ok. 12. tygodnia ciąży wchodzi w mejozę, ale zasadniczo większość z nich wchodzi w pierwszy podział mejotyczny pod koniec życia płodowego. Co ciekawe podział ten nie zachodzi do końca, tylko zostaje zatrzymany pod koniec profazy pierwszego podziału mejotycznego po crossing over. W tym stanie powstała komórka tzw. oocyt I rzędu może pozostać nawet do ostatniej owulacji dorosłej kobiety w okresie przekwitania.
- Dokończenie pierwszego podziału mejotycznego następuje dopiero przed każdą owulacją. W trakcie cyklu oocyt I rzędu wznawia swój podział mejotyczny, w wyniku którego powstaje oocyt II rzędu oraz ciałko kierunkowe. To kolejna różnica pomiędzy oogenezą a spermatogenezą. W wyniku pierwszego podziału mejotycznego powstaje tylko jedna komórka – oocyt II rzędu, która może ostatecznie stać się gametą żeńską. Dzieje się tak dlatego, że w trakcie mejozy zachodzi nierówny podział cytoplazmy i większa jej część trafia właśnie do oocytu. Druga komórka powstała w podziale to właśnie ciałko kierunkowe, które zostaje wydalone do przestrzeni pomiędzy błoną komórkową oocytu a osłonką przejrzystą.
- Oocyt II rzędu wchodzi w drugi podział mejotyczny, który również zostaje zahamowany – tym razem na etapie metafazy II. I właśnie w takiej postaci zostaje on uwolniony podczas owulacji. Kontynuacja drugiego podziału mejotycznego jest uzależniona od procesu zapłodnienia. Na tym etapie mamy już dojrzały pęcherzyk jajnikowy, czyli pęcherzyk Graafa, w którym właśnie znajduje się oocyt II rzędu zahamowany na etapie metafazy drugiego podziału mejotycznego oraz pierwsze ciałko kierunkowe.
- W trakcie owulacji uwolniony z pęcherzyka oocyt II rzędu wraz z otaczającymi go komórkami pęcherzykowymi i ciałkiem kierunkowym zostaje wyrzucony przez skurcz pęcherzyka i skierowany do lejka jajowodu. Dokończenie drugiego podziału mejotycznego zachodzi tylko po zapłodnieniu. Po wniknięciu plemnika efektem zakończenia drugiego podziału mejotycznego jest haploidalny dojrzały oocyt, czyli komórka jajowa, która zawiera prawie całą cytoplazmę oraz ciałko kierunkowe. Wtedy dzieli się również pierwsze ciałko kierunkowe (czyli w wyniku mejozy oogonium powstają w sumie 3 ciałka kierunkowe). Ciałka kierunkowe ulegają zanikowi.
- Około godzinę po zapłodnieniu komórka jajowa i plemnik tworzą diploidalną zygotę, która rozpoczyna podziały mitotyczne prowadzące do rozwoju nowego organizmu.
- Jeżeli nie dojdzie do zapłodnienia oocyt II rzędu zanika.
- W procesie oogenezy w wyniku mejozy pojedynczego oogonium powstaje 1 komórka jajowa i 3 ciałka kierunkowe.
- W przeciwieństwie do spermatogenezy, która trwa przez całe dorosłe życie mężczyzny, oogeneza, pierwsza faza oogenezy zostaje zakończona jeszcze przed narodzinami i zostaje wstrzymana do momentu owulacji u kobiety.
- Żeński noworodek w trakcie narodzin ma 1-2 mln oocytów I rzędu, które znajdują się w pierwotnych pęcherzykach jajnikowych. Następnie w wieku dziecięcym większość z nich zanika. Ostatecznie w okresie pokwitania w jajnikach znajduje się od 20 do 400 tys. oocytów I rzędu. Z tego ok. 420-480 przechodzi procesy różnicowania i dojrzewania i ma szansę przejść owulację na przestrzeni życia kobiety. W trakcie każdego cyklu w jajnikach dojrzewa kilkanaście (15-20) pęcherzyków jajnikowych, ale tylko jeden z nich zostaje uwolniony podczas owulacji. Pozostałe ulegają zanikowi.